


 Каширина Алена, 9 класс, ГУ "Лесная общеобразовательная средняя школа Железинского района", Казахстан, Палодарская область, Железинский район, с.Лесное
Каширина Алена, 9 класс, ГУ "Лесная общеобразовательная средняя школа Железинского района", Казахстан, Палодарская область, Железинский район, с.Лесное
 Научный руководитель: Панова Снежана Алексеевна, учитель биологии и химии, ГУ "Лесная общеобразовательная средняя школа Железинского района", Казахстан, Палодарская область, Железинский район, с.Лесное
Научный руководитель: Панова Снежана Алексеевна, учитель биологии и химии, ГУ "Лесная общеобразовательная средняя школа Железинского района", Казахстан, Палодарская область, Железинский район, с.Лесное
Доклад к презентации Генетика кошек Каширина Алёна 9 класс.docx (21 КБ)
Приложение 2 Сборник задач по генетике кошек.docx (97 КБ)
Проектно-исследовательская работа Генетика кошек.docx (77 КБ)
Презентация Генетика кошек Каширина А., 9 кл.ppt (836 КБ)
Приложение 1 Узнай гены своей кошки.docx (1 024 КБ)
Глоссарий
В данной проектной работе использовались следующие термины:
Генетика – наука, изучающая закономерности наследственности и изменчивости, а также биологические механизмы, их обеспечивающие.
Ген – наименьший участок хромосомы, обусловливающий синтез определенного фермента. Основная единица наследственности для данного признака.
Аллели - альтернативные формы одного и того же гена, определяющие альтернативные признаки.
Локус - местоположение аллеля в хромосоме.
Гомозигота - несет согласованную пару генов для определенной характеристики, которая может быть как в доминантном, так и в рецессивном состоянии: AA или aa.
Гетерозигота - несет две различных характеристики в паре генов. Обычно один доминантный и один рецессивный ген: Аа.
Генотип - генетическая информация, заложенная в организме, комбинация всех аллелей индивидуума. Генотип как правило не соответствует внешнему проявлению.
Фенотип - есть внешнее выражение генотипа, но не выявляет весь генотип.
Доминантный аллель - аллель, определяющий фенотип как в гомозиготном, так и в гетерозиготном состоянии.
Рецессивный аллель - аллель, определяющий фенотип только в гомозиготном состоянии.
Введение
Первое свидетельство об одомашнивании кошек, относится к 2500 году до н. э. и получено из Египта, где были обнаружены около сотни тысяч их мумифицированных останков.
В Китае этих животных знали за тысячу лет до нашей эры, в Японии о них узнали немного позже. Изображения кошек можно увидеть на рисунках из Египта и Египтяне обожествляли этих животных и считали, что они обладают магическими свойствами.
К концу XVIII в Европе у кошек появилась репутация регулятора численности грызунов. Это способствовало их распространению как необходимых домашних животных.
Однако интерес к кошкам, как к экспонатам выставок, пробудился лишь в конце XIX века. Люди стали отмечать различия в окраске их шерсти и ее рисунках. А с зарождением и развитием генетической науки специалисты по разведению кошек стали интересоваться созданием новых пород и совершенствованием уже существующих.
Кошки не могут быть традиционным объектом генетических исследований. Они поздно становятся половозрелыми, плодовитость их невысока, содержание этих животных в лабораторных условиях требует больших материальных затрат. С другой стороны, широчайшая изменчивость собак и кошек и "традиционность" их как физиологических объектов способствовали тому, что целый ряд ученых посвятили себя изучению их генетики.
Исследования генома кошки важны и для человеческой медицины - кошки имеют не менее 250 наследственных заболеваний, общих с человеком и поражаются зачастую аналогичными инфекциями, что и человек. Домашняя кошка предоставляет отличную модель для изучения инфекционных заболеваний, в том числе модель для изучения человеческого СПИДа.
Начало исследований генетики кошки относится к 20-м годам XX века.
В настоящее время проблемы генетики кошек привлекают все большее и большее внимание ученых нашей страны. На сегодняшний день большинство руководств по генетике кошек написаны специалистами по сельскохозяйственным животным или молекулярной генетике, которые недостаточно хорошо ориентируются в фелинологии в целом и часто слишком обобщенно судят о частной генетике животных этих видов. Кроме того, часто статьи иностранных авторов, переводятся без учета особенностей фелинологической терминологии, принятой у нас в стране. Иногда представленные данные также не согласуются с принятой в России генетической номенклатурой.
В то же время, несмотря на обилие фелинологической литературы, практически нет руководств по генетике отдельных пород, описывающих их специфику и селекционно-значимые признаки. Особенно это касается молодых пород, к которым относятся и британские кошки. Использующие такую литературу заводчики, по сути, являются любителями, а не профессионалами-селекционерами. Незнание генетики своего объекта ведет к повышению уровня некомпетентности фелинологов.
Одним из важнейших селекционных признаков для кошек, является их окрас. Знания по типированию - определению вероятного генотипа животного по окраске и текстуре шерсти - позволяют проводить анализ наследования селекционно-важных признаков у скрещивающихся британских короткошерстных и длинношерстных домашних кошек.
Таким образом, на современном этапе развития фелинологии большое значение приобретают точные методы определения окраса кошки. В племенной работе с кошками часто встречаются случаи ошибочного определения окраса при осмотре котят методистами племенных комиссий клубов любителей кошек или выставочной оценке взрослых животных, что влечет за собой неадекватную оценку генотипа и сомнения в генетической чистоте производителей. Наиболее точное определение генотипа родителей позволяет правильно производить отбор и подбор пар по такому важному в разведении кошек признаку, как окраска шерсти. Встает вопрос о влиянии генов, определяющих окраску, длину и структуру шерсти, на морфологию волоса не только на макро-, но и на микроскопическом уровне. Для чего необходимо уточнить строение волоса кошки, поскольку имеющиеся в литературе данные разрозненные и неполные.
Другая проблема, связана с тем, что большинство заводчиков не рассматривают окрас шерсти кошки, как основной признак породы, но все же, окраска шерсти для них селекционно-значима, в целях получения котят желаемого окраса. Для скрещивания часто используют кошек с различными окрасами, следовательно, кошки из питомника далеко не всегда имеют точно определенный генотип относительно генов окраса.
Объектом исследования является генетическое разнообразие кошек села Лесное.
Предметом исследования являются коты и кошки жителей села Лесное.
Цель: определение генетического разнообразия кошек села Лесное.
Для достижения цели были поставлены следующие задачи:
Гипотеза: по окраске кошки можно определить наличие и отсутствие у нее тех или иных генов.
Методы исследования: описание, наблюдение, поисковый метод, метод анализа.
Результаты исследования:
Некоторые гены изучены и их расположение известно. Эти гены сгруппированы по оказываемому ими эффекту: гены, отвечающие за форму тела; за текстуру и и длину шерсти; за окрас и рисунок шерсти.
Гены окраса делятся на три группы: гены цвета - контролируют цвет шерсти и ее плотность; гены рисунка - рисунок и интенсивность цвета; и гены маски - отвечают за тип и степень проявления маски на основном окрасе (пример маски - сиамское пятно на голове).
Эти гены формируют основные внешние признаки: уши, хвост и лапы. Этих генов тысячи, но пока определены только некоторые:
Ген вислоухости (Scottish-fold gene): естественная аллель "fd" рецессивна и формирует обычные уши. Мутация "Fd" доминирует и приводит к прижатым к голове ушам у Шотландской Вислоухой кошки. Мутация опасна для гомозиготов, поэтому скотишей не спаривают между собой.
Ген Японского бобтейла (Japanese Bobtail gene): естественная аллель "Jb" доминирует и формирует хвост нормальной длинны. Мутация "jb" рецессивна и формирует короткий хвост у Японских Бобтейлов. В отличие от мутации Манкской кошки, эта мутация не приводит к деформации позвоночника.
Ген Манкской кошки (Manx gene): естественная аллель "m" рецессивна и формирует хвост нормальной длинны правильное строение позвоночника. Мутация "M" доминантна и приводитк к отсутствию хвоста и укорочению позвоночника. Такая мутация летальна для гомозиготов. Для гетерозиготов это часто приводит к разным опасным болезням типа недержания, запоров и т.п.
Ген полидактилии (polydactyl gene): естественная аллель "pd" рецессивна и формирует правильное количество пальцев. Мутация "Pd" доминаннтна и приводит к образованию лишних пальцев, чаще на передних лапах. Интересно, что у человека тоже обнаружен подобный ген. Гомозиготные люди с шестью пальцами на каждой руке передают такую особенность своим детям, гетерозиготные - каждому четвертому ребенку, даже с нормальной матерью: ген доминантный. Несмотря на это, у людей обычно пять пальцев. Если мутация не благоприятна для выживания или как-то ухудшает возможность продолжения рода, она никогда не охватывает большинство представителей данного вида (как людей, так и кошек).
1.1.2 Гены строения шерсти
Эти гены отвечают за длину и текстуру шерсти.
Гены Сфинкса (Sphinx gene) определяют будет кошка лысой или нет. Естественная аллель "Hr" доминантна и формирует обычную шерсть. Мутация "hr" рецессивна и приводит к бесшерстности (или почти бесшерстности) Сфинксов.
Ген длинношерстности (longhaired gene) определяет короткая или длинная будет шерсть. Естественная аллель "L" доминантна и формирует короткую шерсть. Мутация "l" рецессивна и образует длинную шерсть Персидской, Ангорской, Сибирской и других кошек.
Ген Корниш Рекса (Cornish Rex gene) определяет прямая или кудрявая будет шерсть. Естественная аллель "R" доминантна и формирует прямые волосы шерсти. Мутация "r" рецессивна и приводит к очень короткой кудрявой шерсти у Корниш Рексов.
Ген Девон Рексов (Devon Rex gene) определяет прямая или кудрявая будет шерсть. Естественная аллель "Re" доминантна и формирует прямые волосы шерсти. Мутация "re" рецессивна и приводит к очень короткой кудрявой шерсти Девон Рексов. В отличие от Корнишей, у шерсти Девон Рексов есть жесткие волосы.
Ген Орегонского Рекса (Oregon Rex gene) определяет прямая или кудрявая будет шерсть. Естественная аллель "Ro" доминантна и формирует прямые волосы шерсти. Мутация "ro" рецессивна и приводит к очень короткой кудрявой шерсти у Орегонского Рекса. Как и у Корнишей отсутствуют жесткие волосы.
Обратите внимание, что хотя три разные мутации приводят к одинаковому результату, вызывают их три различных гена.
Ген Американской Жесткошерстной Кошки (American Wirehair) определяет жесткость шерсти. Естественная аллель "wh" рецессивна и формирует обычную мягкую шерсть. Мутация "Wh" доминантна и приводит к образованию короткой упругой, ''проволочной'' шерсти у Американской Жесткошерстной Кошки.
Эти гены отвечают за цвет, рисунок и их насыщенность у шерсти. У кошек это наиболее важная отличительная черта, внешне намного ярче проявляющаяся, чем другие.
Гены этой группы подразделяют на три подгруппы.
Гены цвета шерсти
Гены черного цвета шерсти
Эти гены контролируют цвет шерсти. Задействованы три аллели - черного, темно-коричневого и светло-коричневого цвета. Известна также аллель альбинизма.
Аллель черного цвета "B" доминантна и формирует черную и черно-коричневую табби шерсть, что зависит от гена агути. На самом деле, черный - это очень темный коричневый, настоящий черный теоретически невозможен.
Аллель темно-коричневого цвета "b" - рецессивна черному, но доминантна к светло-коричневому и формирует шерсть от черной до коричневой.
Аллель светло-коричневого цвета "bl" рецессивна и к черному, и к темно-коричневому. Формирует цвет от черного до средне-коричневого.
Гены оранжевого окраса шерсти
Аллель отсутствия оранжевого "o" приводит к явновыраженному черному или коричневому цвету шерсти.
Аллель оранжевого "o" - мутация, преобразует черный или коричневый окрас в рыжий и подавляет эффект не-агути гена (все рыжие кошки - табби).
Эти гены полозависимы - они расположены в "X" хромосоме и не зависят от хромосомы "Y". Поэтому у коты могут быть или рыжими или нет, т.к. ''X'' хромосома у них одна, и если это аллель не-оранжевого - "o", то будет представлен основной цвет (черный, темно-коричневый или светло-коричневый). Если же это аллель "O", то цвет шерсти будет рыжим.
У кошек две хромосомы "X", поэтому вариантов гораздо больше. Если кошка гомозиготна по не-оранжевой аллели - "oo", шерсть может быть любого основного цвета. Если гомозиготна по оранжевой аллели - "OO", то все основные цвета переходят в рыжий. Если кошка гетерозигона - "Oo", то получаются очень элегантные окрасы - черно-оранжевые черепаховый или полосатый.
Сразу после оплодотворения, когда клетка еще строится, активизируется процесс ''рационализации'' всех полозависимых характеристик, включая гены оранжевого. В этом специфическом процессе в каждой клетке случайным образом подавляется один из генов оранжевого (видимо, под влиянием остальных генов). Каждая клетка несет, таким образом, только один ген оранжевого.
Так как в процессе рационализации участвует множество клеток, только несколько из них в конечном счете определяют цвет шерсти (гены других клеток игнорируются). Если определяющие зиготы были гомозиготны для не-оранжевого - "oo", то все клетки будут содержать "o", и шерсть будет не-оранжевой. Аналогично, если зиготы были гомозиготны для оранжевого - "OO", то все клетки будут содержать "O", и шерсть будет рыжей. Но если зиготы гетерозиготны - "Oo", то часть клеток будет содержать "O", а часть - "o". В этом случае, часть шерсть будет рыжей, часть не-рыжей - вот он, черепаховый окрас!
У котят-кошек по две "X" хромосомы и, следовательно, по два гена оранжевого, от каждого из родителей. Приняв ради простоты равновероятное наследование любой аллели от каждого родителя (что конечно не так), получим, что четверть всех кошек будут не-оранжевыми, четверть - оранжевыми, и половина - черепаховыми.
Котята-коты, наоборот, имеют только одну "X" хромосому и, следовательно один ген оранжевого. При равновероятном наследовании генов от родителей, половина котят будет не-оранжевыми, а вторая половина - оранжевыми. Получается, что рыжих котов должно быть вдвое больше, чем кошек.
Наше допущение, однако, не правильное. Ген оранжевого размещается близко к центромере и часто повреждается во время мейоза. Эти повреждения способствуют к замене оранжевой аллели на не-оранжевую, что уменьшает вероятность наследования оранжевого гена, примерно до 3:7. Поэтому среди кошек не-оранжевых 49%, черепаховых 42% и только 9% рыжих. Среди котов соотношение оранжевых к не-оранжевым - 70% yf 30%, то есть в три раза больше, чем у кошек!
Зато черепаховых котов не бывает. Исключение составляют коты-гермафродиты с генетической структурой "XXY". Они могут быть черепаховыми, т.к. у них две "X" хромосомы, но такие коты стерильны. Хотя они имеют все внешние половые признаки котов, фактически они являются недоделанными кошками и рожать не могут.
Гены насыщенности цвета шерсти
Эти гены контролируют однородность пигмента в шерсти и существуют в двух аллелях: плотный - "D" и разбавленный - "d".
Естественная аллель "D" доминантна и располагает пигмент равномерно по каждому волосу, делая цвет шерсти глубоким и чистым. Основными цветами являются черный, темно- и средне-коричневый и оранжевый.
Аллель разбавленного "d" - рецессивная мутация, располагает пигмент микроскопическими группами окруженными обесцвеченными участками. Разбавленными цветами могут быть голубой, желтовато-коричневый, бежевый, или кремовый.
Гены альбиносности
Первым из генов, влияющих на гены цвета шерсти проявлением рисунка является ген альбиносности. Этот ген контролирует количество окрашенной поверхности тела и существует в пяти аллелях:сплошной окрас - "C", Бурма (Burmese) - "cb", Сиамский (Siamese) - "cs", альбинос с голубыми глазами (blue-eyed albino) - "ca", и альбинос (albino) - "c".
Доминантным является естественная аллель сплошного окраса - "C". Еее иногда называют аллелью не-альбиносности.
Аллель Бурмы (Burmese) - "cb" - мутация, рецессивна к аллели сплошного окраса, равноценна сиамскому и доминантна к остальным аллелям. Приводит к слабовыраженному альбинизму, осветляя черный до темно-коричневого, называемого собольим (sable) у Бурмезских кошек, и к появлению зеленых или золотисто-зеленых глаз.
Аллель Сиамского окраса - мутация, рецессивна к аллели сплошного окраса, равноценна Бурмезскому окрасу и доминантна к остальным аллелям. Приводит к среднему альбинизму, осветляя основной цвет от черного или коричневого до светло бежевого, с темно-коричневым пятном классического сиамского рисунка, и к появлению голубого окраса глаз.
Аллели Бурманского и Сиамского окраса имеют одинаковую доминантность. Это делает возможным, при наследовании обоих аллелей - "cbcs", появление Сиамского рисунка на темном теле и зеленовато-голубых (аквамариновах) глаз - Тонкийский окрас (Tonkinese).
Аллели альбиноса с голубыми глазами - "ca", мутация доминантная к аллели альбиноса и рецессивная к остальным. Выглядит почти как полный альбинос, с очень бледным, просвечивающим через белый цвет основным цветом и светло-голубыми глазами.
Аллель альбиноса - "c", мутация рецессивная к остальным. Приводит к полностью прозрачно-белой шерсти и розовым глазам.
Ген Агути
Следующим геном, контролирующим окрас, является ген Агути (agouti). Этот ген создает тиккинг шерсти и существует в двух аллелях: Агути - "A" и не-агути - "a".
Доминантной является естественная аллель агути - "A". Она образует к образованию на каждом волосе полосок (тиккинга), что приводит к образованию рисунка табби.
Аллель не-агути "a" - рецессивная мутация, подавляющая тиккинг, что приводит к образованию сплошного окраса шерсти. Этот ген проявляет себя в сочетании с геном не-оранжевого, образуя сплошной черный, темно-коричневый или светло-коричневый окрасы, а в сочетании с геном оранжевого - подавляя оранжевый цвет в окрасе шерсти.
Ген Табби
Последним из группы генов рисунка является ген табби (tabby). Этот ген существует в аллелях:тигровый или полосатый (mackerel, striped) табби - "T", Абиссинский (Abyssinian) - "Ta", пятнистый или классический (blotched) табби - "tsp" , мраморный - "tb".
Естественная аллель тигрового табби - "T", равнодоминантна с аллелью Абиссинского табби и доминантна к аллели классического табби. Образует полосатый рисунок, с вертикальными, не-агути полосками на агути окрасе остальной шерсти. Это наиболее типичный рисунок, выработавшийся для маскировки на травянистых равнинах, с длинными и прямыми тенями.
Пятнистый табби - это генетически полосатый табби, но с прерывистыми полосками из-за полигенных взаимодействий. Специфического гена пятнистости не существует. Такой пятнистый рисунок выработался для маскировки в условиях леса, где тени от солнечного света, пробивающегося через листву, ложатся пятнами. Не надо, однако, путать пятна наших кошек с розетками истинно пятнистых кошаек, их формируют совершенно другие гены.
Аллель Абиссинского окраса - "Ta", мутация равнодоминантная с тигровым табби и доминантная к аллели классического табби. Образует шерсть сплошного агути окраса, без полос или пятен. Такой окрас типичен для животных обитающих в пустынной местности.
Интересен случай, когда у кошки присутствуют гены и тигрового, и абиссинского окраса - "TTa". Это приводит к уникальному рисунку шерсти - общий бежевый фон и каждый волос типпирован явно выраженным цветом. Бридерам удается получить и мягко-золотистый фон (золотистая шиншилла).
Аллель пятнистого или классического окраса - "tb", рецессивна к двум предыдущим. Образует неправильные не-агути пятна или разводы на агути фоне. Если рисунок симметрично расположен по бокам - получается очень красивый окрас.
Гены интенсивности цвета
Первый из генов влияющих на интенсивность цвета шерсти является ген подавляющий цвет (ингибитор). Он существует в двух аллелях: не-ингибитор - "i", и ингибитор - " I ".
Естественная аллель не-ингибитор "i" рецессивна. Образует шерсть с волосками, прокрашенными по всей длине.
Аллель ингибитор "I" - доминантная мутация и выражается в появлении не прокрашенных участков волоса.
Аллель ингибитор может проявляться в разной степени. В слабовыраженном случае не окрашены только короткие участки у основания волоса, до четверти длины - такой окрас называют дымчатым(smoked). Средне выраженный вариант, когда волос не окрашен до половины, окрас называется затушеванным (shaded). При полном проявлении волосы не окрашены на три четверти. Этот окрас называется шиншиллой (chinchilla) или типпированным.
Ни одна из аллелей не меняет цвета и рисунка шерсти.
Ген пятнистости (Spotting Gene)
Следующим геном, контролирующим насыщенность цвета, является ген белых пятен. Этот ген отвечает за наличие и конфигурацию белых пятен на основном рисунке шерсти. Ген существует в четырех аллелях: беспятнистый - "s", пятнистый -"S", частично-пятнистый - "Sp", и бирманский - "sb". Существование двух последних часто подвергается сомнению - выделить их проявление очень трудно.
Естественной является аллель беспятнистого - "s". Она рецессивна, образует шерсть без белых пятен.
Аллель пятнистого - "S", доминантная мутация. Приводит к появлению белых пятен на различных участках тела. Проявляется ата аллель по-разному - от маленького белого пятнышка у черных кошек, до полностью белых с несколькими черными волосками.
Аллель частично-пятнистого - "Sp", если она все же существует - это разновидность аллели пятнистого. Классический рисунок этой аллели - перевернутая белая буква "V" с вершиной посередине лба, расходящаяся к глазам. The подбородок, живот, лапы - белые. В зависимости от степени проявления этой аллели, количество белого может уменьшаться до белого медальона или пятнышка на лбу.
Аллель бирмы - "Sb", если и она существует - это также разновидность аллели пятнистого, приводящая к белым ногам. В зависимости от степени проявления этой аллели, количество белого может уменьшаться так, что белыми остаются только ''тапочки'' на ступнях.
В отличие от гена белого и гена альбиносности, ген белого пятна не влияет на цвет глаз: если у белой кошки зеленые глаза, это значит, что у кошки одно белое пятно на все тело, а сама она не белая!
Гены доминирующего белого цвета (Dominant-White)
Это последний из группы генов, контролирующих насыщенность цвета. Он будет ли шерсть сплошного белого цвета, или нет. Ген существует в трех аллелях: не-белый - "w", белый - "W", и ван - "Wv". Существование аллели ван пока не доказано, возможно это отдельный ген.
Естественной является аллель не-белого - "w". Она рецессивна и приводит к полному проявлению основного цвета и рисунка шерсти.
Аллель белого "W" - доминирующая мутация, приводит к полностью белой шерсти и оранжевым или бледно-голубым глазам. Голубоглазые белые кошки часто бывают глухими, случается, что и с оранжевыми тоже. Бывает что кошки могут быть с глазами разного цвета - одним голубым, вторым оранжевым. Такие кошки часто глухи на одно ухо (со стороны голубого глаза).
Аллель ван - "Wv", если существует, образует классический ван-рисунок: пятно на макушке головы (часто разделенное пополам тонкой белой линией), на ушах и на хвосте. Размер пятен зависит от степени проявления действия аллели. Бывает, что пятно на голове отсутствует и только уши и (или) хвост окрашены белым.
Важно заметить, что генетически белый - это не цвет, а полное подавление обычно присутствующего пигмента. Гетерозиготные белые кошки могут давать цветных котят, иногда совсем без белого.
1.2. Составление и чтение генетических формул окраса.
После знакомства с генетикой пигментогенеза составление и чтение генетических формул окраса не составит большого труда. Часть генотипа кошки записана, так сказать, на ней.
Возьмем для примера голубокремовую кошку. Ее окрас не белый - следовательно, она гомозиготна по рецессивному аллелю гена W - ww, белые пятна отсутствуют - ss, агути-фактор не выражен – аа.
Далее, нам известно, что голубой окрас — это осветленный черный. Осветленный записывается просто — dd.
Относительно черного можно отметить только один доминантный аллель — В-. Состояние же его гомолога по внешнему виду кошки определить невозможно, она может быть и гомо-, и гетерозиготной. То же касается гена альбинизма — мы видим полное окрашивание — (С-), но ничто не мешает этой кошке в своем генотипе нести аллель с или cs. Поскольку кошка черепаховая, ясно, что она гетерозиготна по гену красного окраса — Оо. Отсутствие признаков "дыма" можно записать как ii или как erer. А вот определить на черепаховой кошке, "теплый" ли у нее тон подшерстка, практически нереально.
Точно так же по ее внешнему виду нельзя судить о том, какой тип рисунка закодирован в ее генотипе (но это не означает, что у кошки нет никаких аллелей в локусе Т или самого этого локуса!). Итак, судя по "внешнему виду кошки, ее генотип должен быть записан как: ааВ-С-ddOosswwii(или erer).
Часть данных о генотипе животного мы можем вывести из его родословной, точнее, установить по внешнему облику родителей. Если мать описываемой кошки была шоколадно-мраморная, а отец кремовый мраморный, то аллельное состояние локусов В и Т можно определить как гетерозиготное - Bb и гомозиготное - tbtb. Но если кто-то из родителей имел тигровый рисунок, то мы не можем записать состояние гена как Ttb, так как не знаем, была ли мать гомозиготной по доминантному аллелю Т и передала ли своей дочери именно этот аллель, а не его рецессивный гомолог. Конечно, если отец или мать кошки имеют белый окрас, то никакой информации о состоянии интересующих нас генов получить невозможно. А вот если при наличии шоколадно-мраморной матери отец кошки в родословной записан как лиловый или черный, то мы, скорее всего, имеем дело с подделкой: от таких родителей кошка никак не могла унаследовать доминантный ген О.
Генетические формулы можно использовать для расчета вероятности получения тех или окрасов при вязках. В последние годы появились целые книги таблиц, где приводятся возможные окрасы потомства при различных цветовых вариантах вязок. Однако любой мало-мальски генетически грамотный заводчик легко проведет эти расчеты и без помощи справочной таблицы. Во-первых, следует определить генотипы родителей (так, как это было сделано выше). Допустим, вышеописанную голубокремовую самку мы хотим повязать шоколадным котом, происходящим от лилового отца и черной матери. Генотип кота: aabbC-DdoY (то есть Y-хромосома) sswwii. При составлении таблицы расчета (а за ее основу взята хорошо нам знакомая решетка Пеннета) можно пренебречь теми генами, аллельное состояние которых одинаково у обоих родителей. У потомков оно, конечно, не изменится. Что же касается тех генов, состояние которых нам неизвестно - С, Т и Вl -то двумя последними в этом конкретном случае также можно пренебречь, поскольку от двух особей без агути-фактора ни тэбби, ни шиншилл получить невозможно. Ген С мы тоже пока не будем включать в таблицу, однако вернемся к его рассмотрению позже. Так же, как и в случае с решеткой Пеннета, по горизонтали размещаем варианты генетического набора, образующиеся в половых клетках одного из родителей, по вертикали - другого. На пересечении записываем генотип котенка. По генетической формуле уже легко прочитать окрас (табл. 1).
Таблица 1
Распределение окрасов в потомстве
|
Гаметы отца |
Гаметы матери |
|||
|
B d O |
b d O |
b d o |
b d o |
|
|
b D Y |
Красный кот |
Красный кот |
Черный кот |
Шоколадный кот |
|
b d Y |
Кремовый кот |
Кремовый кот |
Голубой кот |
Лиловый кот |
|
b D o |
Черная черепаховая кошка |
Шоколадная черепаховая кошка |
Черная кошка |
Шоколадная кошка |
|
b d o |
Голубо-кремовая кошка |
Лилово-кремовая кошка |
Голубая кошка |
Лиловая кошка |
2. Материалы и методика
Формулировка закона: это закон популяционной генетики — в популяции бесконечно большого размера, в которой не действует отбор, не идет мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов, все скрещивания случайны — частоты генотипов по какому-либо гену (в случае если в популяции есть два аллеля этого гена) будут поддерживаться постоянными из поколения в поколение.
Закону Харди-Вайнберга соответствует уравнение:

Где 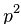 — доля гомозигот по одному из аллелей;
— доля гомозигот по одному из аллелей;  — частота этого аллеля;
— частота этого аллеля; 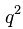 — доля гомозигот по альтернативному аллелю;
— доля гомозигот по альтернативному аллелю;  — частота соответствующего аллеля;
— частота соответствующего аллеля; 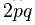 — доля гетерозигот.
— доля гетерозигот.
Пример действия закона при полном доминировании:
Известно, что черная окраска шерсти у кошек определяется генотипом аа. При этом черная окраска может быть или сплошной, или частичной. Генотипы АА и Аа обусловливают все остальное разнообразие типов окраски, но черный цвет при этом полностью отсутствует.
Предположим, что в одной из городских популяций кошек на о. Сахалин из 100 просмотренных животных полную или частичную черную окраску имели 36 животных.
Прямой расчет структуры аллелофонда популяции в этом случае невозможен из-за полного доминирования: гомозиготы АА и гетерозиготы Аа фенотипически неразличимы. Согласно уравнению Харди-Вайнберга частота черных кошек составляет q2 аа. Тогда можно рассчитать частоты аллелей:
q2aa = 36/100 = 0,36; qa = 0,36 –1/2 =0,6; pA = 1 – 0,6 = 0,4
Таким образом, структура аллелофонда данной популяции описывается соотношением: р А + q a = 0,4 + 0,6 = 1. Частота рецессивного аллеля оказалась выше, чем частота доминантного.
Рассчитаем частоты генотипов:
р2 АА = 0,42 = 0,16; 2 pq Аа = 2 ´ 0,4 ´ 0,6 = 0,48; q2aa = 0,62 = 0,36
Однако проверить правильность расчетов в данном случае невозможно, поскольку неизвестны фактические частоты доминантных гомозигот и гетерозигот.
3.Собственные исследования
3.1 Статистика
В рамках исследования проводилась статистика популяции кошек в селе Лесное.
Метод: опрос местных жителей.
На территории села Лесное проживают 112 семей. Из них:
Данный этап исследования проводился с помощью наблюдения и описания фенотипических данных кошек.
Описание фенотипов и дальнейшее определение кариотипов исследуемых объектов были внесены в дневник наблюдений (приложение 1). Приложение 1 Узнай гены своей кошки.docx (1 024 КБ)
Для генов, определяющих окрас волосяного покрова, тип кошки и некоторые другие признаки использовались Стандарты генетической номенклатуры для домашних кошек (СГНДК).
Всего было описано 97 домашних кошек. Из них
В рамках проектной работы мы исследовали только те гены, которые можно было выявить без помощи специалистов. Генетическое разнообразие проводилось по двум типам генов:
Было исследовано три подгруппы генов строения шерсти:
|
1. |
Ген длиношерстности |
Определяет длину шерсти кошек: доминантный аллель – короткошерстная, рецессивный – длинношерстная. |
50 кошек – короткошерстные; 47 – длинношерстных. |
|
2. |
Ген Американской жесткошерстной кошки |
Определяет жесткость шерсти: доминантный аллель - короткая упругая, ''проволочная'' шерсти, рецессивный - обычная мягкая шерсть. |
Все описанные кошки имеют обычную мягкую шерсть. |
|
3. |
Ген Корниш Рекса |
Определяет прямые волосы шерсти: доминантный аллель – прямые волосы шерсти, рецессивный – короткие кудрявые волосы шерсти. |
Все описанные кошки имеют прямые волосы шерсти. |
Было исследовано девять подгрупп генов окраса:
|
1 |
Гены черного цвета шерсти |
Задействованы три аллели: доминантная - черного, рецессивные - темно-коричневого и светло-коричневого цвета. |
60 описанных кошек имеют доминантный аллель, 5 – светло-коричневые, 7 – темно-коричневые. У 25 кошек данный ген отсутствует. |
|
2 |
Гены насыщенности цвета шерсти |
Эти гены контролируют однородность пигмента в шерсти и существуют в двух аллелях: плотный - "D" и разбавленный - "d". |
76 кошек – доминантны по данному гену, 21 – рецессивны. |
|
3 |
Гены Агути |
Доминантный аллель приводит к образованию на каждом волосе полосок (тиккинга), что приводит к образованию рисунка табби. Аллель не-агути "a" - рецессивная мутация, подавляющая тиккинг, что приводит к образованию сплошного окраса шерсти. |
19 кошек доминантны по данному признаку, 78 – рецессивны. |
|
4 |
Гены Тэбби |
Этот ген зависит от гена Агути и существует в четырех аллелях: тигровый или полосатый тэбби - "T", Абиссинский - "Ta" пятнистый или классический тэбби - "tsp" , мраморный - "tb". |
Из 19 кошек доминантных по гену Агути 7 – пятнистых, 5 – Абиссинских (без определенного рисунка) и 7 – тигровых. |
|
5 |
Гены альбиносности |
Этот ген контролирует количество окрашенной поверхности тела и существует в пяти аллелях: сплошной окрас - "C", Бурма - "cb", Сиамский - "cs", альбинос с голубыми глазами - "ca", и альбинос - "c". |
69 кошек имеют сплошной окрас, 28 – окрас Бурма (осветвленный) |
|
6 |
Ген интенсивности цвета |
Проявляется в двух аллелях: ингибитор (доминантный) выражается в появлении не прокрашенных участков волоса, не-ингибитор (рецессивный) образует шерсть с волосками, прокрашенными по всей длине. |
19 кошек доминантны по данному гену, 78 – рецессивны. |
|
7 |
Ген пятнистости |
Этот ген отвечает за наличие и конфигурацию белых пятен на основном рисунке шерсти. Ген существует в четырех аллелях: беспятнистый - "s", пятнистый -"S", частично-пятнистый - "Sp", и бирманский - "sb". |
31 кошка не имеют белых пятен, 66 – доминантны по данному гену |
|
8 |
Гены доминирующего белого цвета |
Он определяет будет ли шерсть сплошного белого цвета, или нет. Ген существует в трех аллелях: не-белый - "w", белый - "W", и ван - "Wv". |
21 кошка доминантна по данному гену, 34 рецессивны, у 42 кошек данный ген отсутствует |
|
9 |
Гены оранжевого окраса шерсти |
Определяет рыжий цвет шерсти и существует в трех аллелях: О_ - рыжий (ОО – кошка, ОУ - кот), о_ - не-рыжий (оо - кошка, оУ- кот), Оо – черепаховый окрас (кошки, реже коты, не имеющие потомства) |
31 кошка доминантна по гену оранжевого окраса шерсти, 66 – рецессивны. Из них 13 рыжих котов, 11 рыжих кошек, 34 не-рыжих котов, 32 не-рыжих кошек, 7 черепаховых кошек. |
С помощью закона Харди-Вайнберга можно определить частоту доминантных и рецессивных гомозиготных особей и гетерозиготных особей в популяции.
|
№ |
Название гена |
Вычисления |
Часто та доми нан тных гомо зигот |
Часто та реце ссив ных гомо зигот |
Часто та гетеро зигот |
|||||||||||||||||
|
1 |
Ген длиношерстности |
q2ll= 47/97=0,485 ql=0,485ᶺ(-1/2)=1,44 pL=1-1,44= - 0,44 p2LL=(-0,44)ᶺ2=0,1936 2pqLl= 2*1,44*(-0,44)=-1,2672 q2ll=1,44ᶺ2=2,0736 Проверка |
0,44 |
2,07 |
1,26 |
|||||||||||||||||
|
q2 |
|
2qp |
|
p2 |
|
|
||||||||||||||||
|
2,0736 |
+ |
-1,2672 |
+ |
0,1936 |
= |
1 |
||||||||||||||||
|
2 |
Ген Американской жесткошерстной кошки |
Все особи рецессивные гомозиготы (whwh), закон Харди-Вайнберга не применим |
|
|
|
|||||||||||||||||
|
3 |
Ген Корниш Рекса |
Все особи доминантные гомозиготы (RR), закон Харди-Вайнберга не применим |
|
|
|
|||||||||||||||||
|
4 |
Гены черного цвета шерсти |
q2вв= 12/72=0,17 qв=0,17ᶺ(-1/2)=2,43 pВ=1-2,43= - 1,43 p2ВВ=(-1,43)ᶺ2=2,04 2pqВв= 2*2,43*(-1,43)=-6,94 q2вв=2,43ᶺ2=5,9 Проверка |
1,43 |
2,43 |
6,94 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
2,04 |
+ |
-6,94 |
+ |
5,9 |
= |
1 |
||||||||||||||||
|
5 |
Гены насыщенности цвета шерсти |
q2dd= 21/97=0,22 qd=0,22ᶺ(-1/2)=2,1 pD=1-2,1= - 1,1 p2DD=(-1,1)ᶺ2=1,21 2pqDd= 2*2,1*(-1,21)=-4,62 q2dd=2,1ᶺ2=4,41 Проверка |
1,1 |
2,1 |
4,62 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
1,21 |
+ |
-4,62 |
+ |
4,41 |
= |
1 |
||||||||||||||||
|
6 |
Гены Агути |
q2аа= 78/97=0,8 qа=0,8ᶺ(-1/2)=1,1 pА=1-1,1= - 0,1 p2АА=(-0,1)ᶺ2=0,01 2pqАа= 2*1,1*(-0,1)=-0,22 q2аа=1,1ᶺ2=1,21 Проверка
|
0,1 |
1,1 |
0,22 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,01 |
+ |
-0,22 |
+ |
1,21 |
= |
1 |
||||||||||||||||
|
7 |
Гены Тэбби |
q2tt= 7/19=0,37 qt=0,37ᶺ(-1/2)=1,65 pT=1-1,65= - 0,65 p2TT=(-0,65)ᶺ2=0,4 2pqTt= 2*1,65*(-0,65)=-2,1 q2tt=1,65ᶺ2=2,7 Проверка
|
0,65 |
1,65 |
2,1 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,4 |
+ |
-2,1 |
+ |
2,7 |
= |
1 |
||||||||||||||||
|
8 |
Гены альбиносности |
q2cc= 28/97=0,289 qc=0,289ᶺ(-1/2)=1,86 pC=1-1,86= - 0,86 p2CC=(-0,86)ᶺ2=0,74 2pqCc= 2*1,86*(-0,86)=-3,2 q2cc=1,86ᶺ2=3,46 Проверка
|
0,86 |
1,86 |
3,2 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,74 |
+ |
-3,2 |
+ |
3,46 |
= |
1 |
||||||||||||||||
|
9 |
Ген интенсивности цвета |
q2ii= 78/97=0,8 qi=0,8ᶺ(-1/2)=1,1 pI=1-1,1= - 0,1 p2II=(-0,1)ᶺ2=0,01 2pqIi= 2*1,1*(-0,1)=-0,22 q2ii=1,1ᶺ2=1,21 Проверка |
0,1 |
1,1 |
0,22 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,01 |
+ |
-0,22 |
+ |
1,21 |
= |
1 |
||||||||||||||||
|
10 |
Ген пятнистости |
q2ss= 31/97=0,32 qs=0,32ᶺ(-1/2)=1,77 pS=1-1,77= - 0,77 p2SS=(-0,77)ᶺ2=0,59 2pqSs= 2*1,77*(-0,77)=-2,72 q2ss=1,77ᶺ2=3,13 Проверка |
0,77 |
1,77 |
3,13 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,59 |
+ |
-2,72 |
+ |
3,13 |
= |
1 |
||||||||||||||||
|
11 |
Гены доминирующего белого цвета |
q2ww= 34/55=0,62 qw=0,62ᶺ(-1/2)=1,27 pW=1-1,27= - 0,27 p2WW=(-0,27)ᶺ2=0,07 2pqWw= 2*1,27*(-0,27)=-0,68 q2ww=1,27ᶺ2=1,61 Проверка |
0,27 |
1,27 |
0,68 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,07 |
+ |
-0,68 |
+ |
1,61 |
= |
1 |
||||||||||||||||
|
12 |
Гены оранжевого окраса шерсти |
q2oo= 31/97=0,32 qo=0,32ᶺ(-1/2)=1,77 pO=1-1,77= - 0,77 p2OO=(-0,77)ᶺ2=0,59 2pqOo= 2*1,77*(-0,77)=-2,72 q2oo=1,77ᶺ2=3,13 Проверка |
0,77 |
1,77 |
3,13 |
|||||||||||||||||
|
p2 |
|
2qp |
|
q2 |
|
|
||||||||||||||||
|
0,59 |
+ |
-2,72 |
+ |
3,13 |
= |
1 |
||||||||||||||||
Заключение
Список используемых источников:
1. http://zcats.ru/biblioteka/otdelnye_publikatsii/416-koshki-v-derevne.html
